" ส่วนแรก
เอนไซม์ที่กระตุ้นขั้นตอนที่สี่ของวงจรเครบส์คือ α-keto กลูตาเรต ดีไฮโดรจีเนส; เอนไซม์นี้เป็นเอนไซม์เชิงซ้อนที่คล้ายกับไพรูเวตดีไฮโดรจีเนสมาก ทั้งสองประกอบด้วยโปรตีน 48-60 ซึ่งมีการรับรู้กิจกรรมของเอนไซม์ที่แตกต่างกันสามอย่างและมีโคแฟกเตอร์ของเอนไซม์เหมือนกัน เป็นเอ็นไซม์ที่คล้ายคลึงกันมากเพราะพวกมันทำปฏิกิริยากับซับสเตรตที่คล้ายคลึงกัน: ทั้งไพรูเวตและแอล "α-คีโตกลูตาเรต คือ α- กรดคีโต กลไกการออกฤทธิ์ของสารเชิงซ้อนของเอนไซม์ทั้งสองจะเหมือนกัน
การโจมตีโดยไทอามีน ไพโรฟอสเฟตบนคาร์บอนิล (C = O) ของ "α-ketoglutarate นำไปสู่การดีคาร์บอกซิเลชันและเกิดอนุพันธ์ของคาร์บอกซีไฮดรอกซีโพรพิล ด้วยการถ่ายโอนในภายหลังไปยัง lipoamide กระบวนการรีดอกซ์ภายในจะเกิดขึ้นจากที่ได้รับ lipoamide carboxy-derivative หรือ succinyl lipoamide
จากนั้น succinyl lipoamide ทำปฏิกิริยากับ coenzyme A เพื่อให้ succinyl coenzyme A (ซึ่งยังคงอยู่ในวงจร krebs) และ lipoamide ที่ลดลงซึ่งถูก reoxidized โดย FAD: FADH2 ที่เกิดขึ้นจะถูก reoxidized โดย NAD + และได้รับ NADH ดังนั้น ในขั้นตอนนี้ การกำจัดคาร์บอนจากโครงกระดูกคาร์บอนเป็นครั้งที่สองจึงเกิดขึ้น ในรูปของคาร์บอนไดออกไซด์
หมู่เอซิลที่เชื่อมโยงกับโคเอ็นไซม์ A อยู่ในรูปแบบกระตุ้น กล่าวคือ มีปริมาณพลังงานสูง ดังนั้นจึงเป็นไปได้ที่จะใช้ประโยชน์จากพลังงานของซัคซินิล โคเอ็นไซม์ เอ

ในขั้นตอนที่ 5 ของวงจรเครบส์ ซัคซินิลโคเอ็นไซม์เอจะอยู่ภายใต้การกระทำของ ซัคซินิล ไธโอคิเนส; มีการตั้งสมมติฐานสองข้อเกี่ยวกับรูปแบบการดำเนินการ: เราจะอธิบายเพียงข้อใดข้อหนึ่งจากสองข้อนี้เนื่องจากเป็นข้อที่ได้รับการรับรองมากที่สุด ตามสมมติฐานนี้ ซัคซินิลโคเอ็นไซม์ A ถูกโจมตีโดยไนโตรเจนของฮิสทิดีน (Hys) ของเอ็นไซม์: โคเอ็นไซม์ A จะถูกปลดปล่อยออกมาและแอดดักต์ที่ได้มาจากฮิสทิดีนจะก่อตัวเป็นตัวกลาง นั่นคือ ซัคซินิล-เอ็นไซม์ (หรือซัคซินิล-ไฮส์) ออร์โธฟอสเฟตทำหน้าที่ในตัวกลางนี้ นำไปสู่การปลดปล่อย ซัคซิเนตและการก่อตัวของฟอสโฟเอนไซม์ ฟอสโนไซม์ ซึ่งถูกโจมตีโดยกัวโนซีน ไดฟอสเฟต (GDP) ทำให้เกิดกวาสโนซีน ไตรฟอสเฟต (GTP) และเอ็นไซม์จะถูกปล่อยออกมา จากมุมมองด้านพลังงาน GTP = ATP: พันธะที่ให้พลังงานเหมือนกันในทั้งสองสปีชีส์ (เป็นพันธะแอนไฮไดรด์ระหว่างฟอสโฟริล Β และฟอสโฟริล γ) ในบางกรณี GTP ถูกใช้เป็นวัสดุที่มีปริมาณพลังงานสูง แต่โดยปกติ GTP จะถูกแปลงเป็น ATP โดยการกระทำของเอนไซม์ นิวคลีโอไซด์ไดฟอสโฟไคเนส (กปปส.); เป็นเอนไซม์ที่พบในเซลล์และกระตุ้นปฏิกิริยาต่อไปนี้:
N1TP + N2DP → N1DP + N2TP
NiTP ® นิวคลีโอไซด์ไตรฟอสเฟตทั่วไป
NiDP ® นิวคลีโอไซด์ไดฟอสเฟตทั่วไป
มันเป็นปฏิกิริยาย้อนกลับ ในกรณีของเรามันเกิดขึ้น:
GTP + ADP → GDP + ATP
ดังนั้นมันสามารถไปทางขวาหรือทางซ้ายได้แม้สำหรับการแปรผันเล็กน้อยในความเข้มข้นของรีเอเจนต์
หากวัฏจักรเครบส์ดำเนินไปด้วยความเร็วจนนำไปสู่การผลิต ATP ที่สูงกว่าความต้องการพลังงาน ก็จะมี ADP ที่ขาดแคลนในขณะที่มี ATP จำนวนมาก: ปฏิกิริยาที่เร่งปฏิกิริยาโดยนิวคลีโอไซด์ไดฟอสโฟไคเนสคือ ไปทางซ้าย ( GTP สะสมถ้านิวคลีโอไซด์ไดฟอสโฟไคเนสไม่มีสารตั้งต้นเพียงพอเช่น ADP) GTP จึงเป็นสัญญาณบ่งบอกถึงความพร้อมของพลังงาน ดังนั้นจึงทำให้วงจรเครบส์ช้าลง
ขั้นตอนที่หกของวงจรเครบส์นำไปสู่การก่อตัวของฟูมาเรตโดยการกระทำของ ซัคซิเนต ดีไฮโดรจีเนส; เอนไซม์นี้ให้ปฏิกิริยา stereospecific เนื่องจากทรานส์ที่ไม่อิ่มตัว (เป็นอัลคีน) มักก่อตัวขึ้นเสมอ กล่าวคือ fumarate (ในขณะที่ cis isomer คือ maleate) พบซัคซิเนต ดีไฮโดรจีเนสที่เยื่อหุ้มไมโทคอนเดรียชั้นใน ในขณะที่เอ็นไซม์อื่นๆ ทั้งหมดของวงจรเครบส์จะกระจัดกระจายไปทั่วไมโตคอนเดรีย
Succinate dehydrogenase มี FAD เป็นปัจจัยร่วม มันถูกยับยั้งโดย oxaloacetate (การยับยั้งการป้อนกลับ) ในขณะที่มันมี succinate และ fumarate เป็นตัวโมดูเลเตอร์เชิงบวก (ตัวกระตุ้น) ตัวกระตุ้นของมัน เรามาลองทำความเข้าใจว่าทำไม โดยการข้ามไปยังขั้นตอนสุดท้ายของวงจรเครบส์ วัฏจักรของ krebs ต้องการพลังงาน ดังนั้น ความเป็นไปได้เพียงอย่างเดียวที่จะได้รับ oxaloacetate จากผู้ป่วยคือความเข้มข้นของผู้ป่วยสูงมาก : malate เป็นหนึ่งในสารที่มีความเข้มข้นสูงสุดในเซลล์ ปฏิกิริยาที่เปลี่ยน malate เป็น oxaloacetate ยังได้รับการสนับสนุนโดย ความจริงที่ว่าความเข้มข้นของ oxaloacetate นั้นต่ำโดยการกระทำของ citrate synthase ปฏิกิริยาที่เร่งปฏิกิริยาโดยซัคซิเนต ดีไฮโดรจีเนสคือปฏิกิริยาการป้อนอาหารด้วยตนเอง และนี่เป็นวิธีเดียวที่จะทำให้การเปลี่ยนแปลงของมาเลตเป็นออกซาโลอะซีเตตเกิดขึ้น
ความเข้มข้นของ mitochondrial malate จะต้องเข้ากันได้กับความเข้มข้นของ cytoplasmic malate: เฉพาะเมื่อความเข้มข้นของ mitochondrial malate สูงจนรับประกันการเปลี่ยนของ malate เป็น oxaloacetate (ใน krebs cycle) จากนั้น malate ก็สามารถนำมาใช้ใน วิธีอื่น (ซึ่งเป็นไซโตพลาสซึม): ในไซโตพลาสซึม มาลาเตสามารถเปลี่ยนเป็นออกซาโลอะซิเตตซึ่งแอสพาเทตสามารถรับได้โดยการกระทำของ GOT (มันคือทรานส์อะมิเนส) หรือกลูโคสผ่านกลูโคนีเจเนซิส
เรากลับไปที่ขั้นตอนที่เจ็ดของวัฏจักรเครบส์ซึ่งถูกเร่งโดยเอนไซม์ ฟูมาราซี: เติมน้ำด้วยวิธี stereospecific เพื่อให้แอล-มาเลต
ในขั้นตอนสุดท้ายของวัฏจักร Krebs ที่เราได้พูดไปแล้ว การกระทำของ มาเลตดีไฮโดรจีเนส. เอนไซม์นี้ใช้โมเลกุล NAD + ในการเร่งปฏิกิริยา
เราจึงได้สรุปรายละเอียดของขั้นตอนต่างๆ ของวงจรเครบส์
วงจรเครบส์สามารถย้อนกลับได้ทั้งหมด
เพื่อเพิ่มความเร็วของวัฏจักรเครบส์ ความเข้มข้นของเมแทบอไลต์ที่มีอยู่ในวัฏจักรนั้นจะเพิ่มขึ้น หนึ่งในกลยุทธ์ในการเพิ่มความเร็วของวัฏจักรเครบส์ประกอบด้วยการเปลี่ยนส่วนของไพรูเวตที่เข้าสู่ไมโทคอนเดรียเป็นออกซาโลอะซิเตต (โดยการกระทำของไพรูเวตคาร์บอกซิเลส) และไม่เปลี่ยนมันทั้งหมดเป็นอะเซทิลโคเอ็นไซม์ A ซึ่งจะทำให้ความเข้มข้นของออกซาโลอะซีเตตเพิ่มขึ้นซึ่ง เป็นเมแทบอไลต์ของวงจรเครบส์ ดังนั้นจึงเพิ่มความเร็วของวงจรทั้งหมด
ในรอบเครบส์ NAD + สามรายการจะถูกแปลงเป็น NADH สามรายการและหนึ่ง FAD เป็น FADH2 และยิ่งไปกว่านั้น ยังได้ GTP: โดยการส่งกำลังรีดิวซ์ที่ได้รับจากวงจรเครบส์ จะผลิต ATP เพิ่มเติม ในห่วงโซ่การหายใจ พลังงานรีดิวซ์จะถูกถ่ายโอนจาก NADH และ FADH2 ไปยังออกซิเจน: การถ่ายโอนนี้เกิดจากชุดของเอนไซม์ที่อยู่บนเยื่อหุ้มไมโตคอนเดรียซึ่งในการกระทำของพวกเขาจะนำไปสู่การผลิต ATP
กระบวนการของระบบทางเดินหายใจเป็นกระบวนการ exergonic และพลังงานที่ปลดปล่อยออกมาถูกใช้เพื่อผลิต ATP จุดประสงค์ของเซลล์คือการใช้ประโยชน์จากกระบวนการ exergonic เพื่อทำให้การสังเคราะห์ ATP เกิดขึ้น สำหรับแต่ละโมเลกุลของ NADH ที่เข้าสู่ระบบทางเดินหายใจ จะได้รับ 2.5 โมเลกุลของ ATP และสำหรับแต่ละโมเลกุลของ FADH2 1.5 ของ ATP ความหลากหลายนี้เกิดจากการที่ FADH2 เข้าสู่ระบบทางเดินหายใจที่ระดับต่ำกว่า NADH
ด้วยพลังลดการเผาผลาญแบบแอโรบิก 30-32 ATP (219-233 kcal / mol) จะได้รับโดยมีประสิทธิภาพประมาณ 33% (ประสิทธิภาพของการเผาผลาญแบบไม่ใช้ออกซิเจนประมาณ 2%)

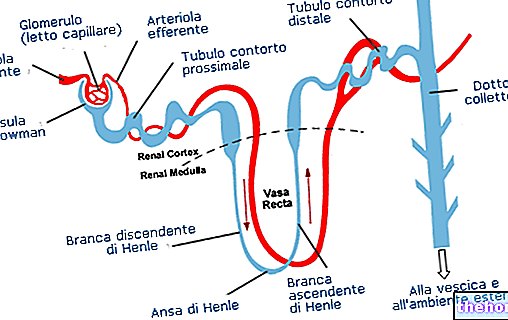


.jpg)








.jpg)



