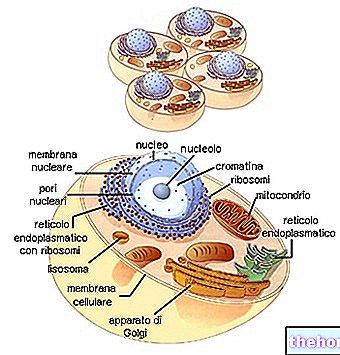
สมาคม (หรือ "การต่อกัน" หรือ "การเชื่อมโยง")
จนถึงปัจจุบัน เราได้ปฏิบัติต่อ Mendelian dihybrid (หรือ polyhybrid) crossing โดยสันนิษฐานว่าคู่ allelic ที่ต่างกันนั้นพบได้จริงในคู่ของโครโมโซมที่คล้ายคลึงกันที่ต่างกัน แต่จำนวนคู่ของโครโมโซมถึงแม้จะแตกต่างกันไปในแต่ละสปีชีส์จะแตกต่างกันไปภายในขอบเขตที่แคบ (ไม่กี่สปีชีส์ ถึงประมาณร้อยโครโมโซม) ในขณะที่จำนวนยีนสามารถนับได้เป็นหมื่น
ว่าตัวละครที่ Mendel เลือกใช้สำหรับการทดลองของเขานั้นแยกออกมาทั้งหมดโดยอิสระ (โดยไม่ทำให้การคำนวณในการแจกแจงฟีโนไทป์ใน F2 ของ polyhybrid cross สับสน) ถือเป็นโอกาสที่ดี หากพบอัลลีลสองคู่ที่ตำแหน่งที่อยู่ติดกัน กฎที่ว่า จะปฏิบัติตามจะเรียกว่ากฎหมายสมาคม
เมื่อรู้ว่าอักขระจำนวนมากมีโลคัสอยู่บนโครโมโซมเดียวและเป็นคู่โครโมโซมที่แยกจากกันอย่างอิสระในไมโอซิส เป็นที่เข้าใจกันดีว่าอักขระสองคู่เกิดขึ้นบ่อยเพียงใด หากสัมพันธ์กับโครโมโซมในผู้ปกครอง สิ่งมีชีวิต ยังคงเกี่ยวข้อง เท่าเทียมกันใน gamete และดังนั้นในสิ่งมีชีวิตที่จะนำสารพันธุกรรมของตัวเอง
ดังนั้นเราจึงเห็นว่า "การเชื่อมโยงเป็นตัวแทนของ" ข้อยกเว้น ห่างไกลจากความเป็นอิสระที่ประกาศไว้ในกฎข้อที่สามของเมนเดล
การแลกเปลี่ยนหรือ "การข้าม" และการรวมตัวใหม่
เมื่อพูดถึงไมโอซิส เราได้ระบุว่ามีสองช่วงเวลาที่แตกต่างกันของการผสมสารพันธุกรรม: ช่วงเวลาหนึ่งคือการแยกโครโมโซมในเซลล์สืบพันธุ์ และนั่นคือสิ่งที่เมนเดลสังเกต
อีกชั่วขณะหนึ่งซึ่งเกิดขึ้นก่อนจริง ๆ คือช่วงเวลาที่โครมาทิดทั้งสี่ของโครโมโซมที่คล้ายคลึงกันแต่ละคู่แลกเปลี่ยนลักษณะที่เหมือนกันหลังจากการแลกเปลี่ยนนี้ปัจจัยสองประการที่เกี่ยวข้องกับโครโมโซมเดียวกันจะเป็นอิสระในเซลล์สืบพันธุ์ การแลกเปลี่ยนเกิดขึ้นเป็นสัดส่วน กับการประมาณครั้งแรก กับความยาวของโครโมโซม และในโครโมโซมที่ยาวกว่านั้นอาจมีมากกว่าการแลกเปลี่ยน
ปรากฏการณ์นี้สามารถตรวจพบได้ทางเซลล์วิทยา โดยการสังเกตไมโอสจำนวนเพียงพอภายใต้กล้องจุลทรรศน์
อัตราการจับคู่ใหม่คืออัตราที่อักขระสองตัวใดๆ ที่เชื่อมโยงในการสร้างโดยผู้ปกครองจะรวมกันอีกครั้งใน F2
หากตำแหน่งทั้งสองอยู่ติดกันโดยสิ้นเชิง ความน่าจะเป็นที่จุดตัดแยกจากกันนั้นแทบจะเป็นศูนย์ อัตราการรวมตัวกันใหม่จะเป็น: n ° recombinants ถ้าตำแหน่งสองตำแหน่งอยู่บนโครโมโซมที่แตกต่างกัน 2 ตัว อัตราการรวมตัวกันใหม่จะเท่ากับ 0.5 (ความน่าจะเป็นเท่ากันสำหรับอักขระสองตัวที่เข้าร่วมในรุ่น P เพื่อสุ่มพบว่าตัวเองอยู่ด้วยกันใน F2) อัตราการรวมตัวกันใหม่จึงสามารถเปลี่ยนแปลงได้ระหว่าง 0.0 ถึง 0.5 สำหรับระยะทางเล็ก ๆ บนโครโมโซม ระยะทางและอัตราการรวมตัวกันใหม่จะเป็นสัดส่วนโดยตรง สำหรับระยะทางที่ไกลกว่า มีความเป็นไปได้ที่การแลกเปลี่ยนสองครั้งเกิดขึ้นระหว่างสองตำแหน่ง ตอนนี้จะเห็นได้ชัดว่าปัจจัยสองประการที่แยกจากกันโดยการแลกเปลี่ยนสองครั้งนั้นเชื่อมโยงกันอีกครั้ง ณ จุดนี้เป็นที่ชัดเจนว่าสัดส่วนระหว่างระยะห่างของตำแหน่งและความน่าจะเป็นของการรวมตัวใหม่จะหายไป
Loci ที่พบว่าเกี่ยวข้องกับโครโมโซมเดียวกันประกอบด้วย 'กลุ่มสัมพันธ์' ตำแหน่งที่อยู่ห่างไกลมากอาจมีความเป็นไปได้ที่จะแยกจากกันโดยการแลกเปลี่ยนซึ่งพวกมันทำตัวเป็นอิสระ แต่แต่ละตำแหน่งจะสัมพันธ์กับอัตราการรวมตัวกันที่ต่ำกว่ากับตำแหน่งกลาง
เมื่อทราบอัตราการรวมตัวกันใหม่ระหว่างยีนหลายคู่ภายในกลุ่มความสัมพันธ์ การสร้าง 'แผนที่พันธุกรรม' สามารถเริ่มต้นได้ โปรดจำไว้ว่าระยะห่างระหว่างสองยีน (a และ b) นั้นแสดงโดยอัตราการรวมตัวกันใหม่ และระยะห่างของ a จากยีนที่สาม c อาจเป็นผลรวมหรือส่วนต่างในแง่ของระยะห่างจาก b ก็ได้ เพื่อสร้างแผนที่ระยะทางซึ่งกันและกัน ซึ่งจะเป็นแผนที่ทางพันธุกรรมภายในกลุ่มของความสัมพันธ์นั้น นั่นคือ ของโครโมโซมนั้น
ตอนนี้เราต้องพิจารณาโดยทั่วไปแนวคิดบางอย่างที่จำกัดการแสดงลักษณะทางฟีโนไทป์ของอักขระจีโนไทป์
ก่อนอื่นเราจะพูดถึงแนวความคิดของการแทรกซึมและการแสดงออก จากนั้นเราจะให้ความสนใจเป็นพิเศษกับปรากฏการณ์ของการควบคุมการกระทำของยีน
ทะลุทะลวง
การแทรกซึมของยีนแสดงถึงความสามารถในการแสดงออกในฟีโนไทป์ การแทรกซึมถูกวัดทางสถิติโดยการนับความถี่ของฟีโนไทป์ที่แสดงอักขระนั้นจาก 100 จีโนไทป์ที่มีอยู่ ลักษณะที่มีการเจาะทะลุ 0.7 เป็นลักษณะที่เกิดขึ้นทางฟีโนไทป์ใน 70% ของความถี่จีโนไทป์
การแสดงออก
การแสดงออกคือการประเมินเชิงปริมาณของระดับของการแสดงออกทางฟีโนไทป์
กฎระเบียบของการกระทำของยีน
เซลล์ผลิตเอนไซม์และโปรตีนทั้งหมดด้วยความเร็วเท่ากันและในเวลาเดียวกัน ยกตัวอย่างเช่น เซลล์ Escherichia coli สามารถให้พลังงานและอะตอมของคาร์บอนจากแลคโตสไดแซ็กคาไรด์ได้เนื่องจากสามารถแตกตัวเป็นกลูโคสและกาแลคโตสได้ด้วยเอนไซม์เบต้ากาแลคโตซิเดส ใน E. coli ปกติที่อาจมีแลคโตส มีประมาณ 3 000 โมเลกุลของเบต้ากาแลคโตซิเดส เท่ากับ 3% ของโปรตีนของเซลล์นั้น ในกรณีที่ไม่มีแลคโตสจะมีเพียงหนึ่งโมเลกุลของเบต้ากาแลคโตซิเดสต่อเซลล์แบคทีเรีย กาแลคโตซิเดสจะถูกสังเคราะห์จากโมเลกุล mRNA ใหม่เมื่อสามารถใช้ได้ สายพันธุ์กลายพันธุ์ของ E. coli ที่อุดมไปด้วยเอ็นไซม์เป็นที่รู้จักกันแม้ในขณะที่ไม่มีแลคโตส: การกลายพันธุ์เหล่านี้เสียเปรียบเมื่อเทียบกับเซลล์ปกติ เนื่องจากพวกมันถูกบังคับให้ใช้พลังงานและวัสดุโดยไม่จำเป็นเพื่อผลิตเอ็นไซม์ซึ่งจะยังคงอยู่โดยไม่มีสารตั้งต้น สารที่ทำให้ปริมาณเอนไซม์เพิ่มขึ้นเช่นเดียวกับแลคโตสจะเรียกว่าสารกระตุ้นในขณะที่เอนไซม์จะเรียกว่าเหนี่ยวนำได้ สารอื่น ๆ ทำให้เกิดการผลิตเอนไซม์บางชนิดในลักษณะเฉพาะเช่นกัน ตัวอย่างเช่นใน E. coli สามารถสร้างกรดอะมิโนทั้งหมดที่มีคาร์บอนและแอมโมเนียม (NH3) การมีอยู่ในตัวกลางในการเพาะเลี้ยงของกรดอะมิโนบางชนิด (เช่นฮิสติดีน) ขัดขวางการผลิตเอนไซม์ทั้งหมดที่เกี่ยวข้องกับ การสังเคราะห์ทางชีวภาพของกรดอะมิโนเอง: จะมีการกล่าวถึงเอ็นไซม์เหล่านี้ว่าสามารถกดทับได้ ในเซลล์แบคทีเรีย โมเลกุล mRNA จะถูกทำลายหลังจากการก่อตัวของพวกมันไม่นาน ด้วยเหตุนี้ การควบคุมการผลิต mRNA จึงหมายถึงการควบคุมการสังเคราะห์ของเอนไซม์ในเวลาเดียวกัน เวลา.
โอเปร่า
เพื่ออธิบายว่าเซลล์แบคทีเรียสามารถควบคุมการผลิตเอ็นไซม์ของตัวเองได้อย่างไร เจคอบและโมโนดได้กำหนดสมมติฐานของโอเปอรอน โอเปอเรเตอร์ถูกสร้างขึ้นโดยยีนหลายตัวที่เกี่ยวข้องกับหน้าที่และเรียงตัวกันโดยไม่ขาดตอนตลอดแนวดีเอ็นเอที่ยืดยาว โอเปอรอนประกอบด้วยยีนที่แตกต่างกันสามประเภท: โปรโมเตอร์ซึ่งการก่อตัวของ mRNA เริ่มต้นขึ้น โอเปอเรเตอร์ ที่ใช้การควบคุม ยีนโครงสร้างตั้งแต่หนึ่งตัวขึ้นไปซึ่งเขียนรหัสสำหรับเอ็นไซม์หรือโปรตีนอื่น ๆ ในระบบเบต้ากาแลคโตซิเดส โอเปอรอนรวมถึงเบตากาแลคโตซิเดสยังมียีนอีกสองตัวที่เข้ารหัสโครงสร้างสำหรับ เอนไซม์อื่นๆ ที่เกี่ยวข้องกับการเผาผลาญแลคโตส ยีนเหล่านี้อยู่ติดกันและถูกถ่ายทอดออกมาทีละส่วนตามเกลียวของ DNA เดียวกันให้เป็นโมเลกุล mRNA ตัวเดียว โมเลกุล mRNA ที่ผลิตขึ้นจึงมีการทำงานในระยะเวลาสั้น ๆ หลังจากนั้นจะถูกทำลายโดยเอนไซม์จำเพาะ
กิจกรรมของโอเปอรอนถูกควบคุมโดยยีนอื่น ตัวควบคุม ซึ่งอาจอยู่ห่างจากโอเปอรอนด้วย: ตัวควบคุมนี้เข้ารหัสโปรตีนที่เรียกว่า รีเพรสเซอร์ ซึ่งดูเหมือนว่าจะจับกับ DNA ที่ยีนโอเปอเรเตอร์ ระหว่างโปรโมเตอร์และยีนโครงสร้างขัดขวางการผลิต mRNA
ในทางกลับกันตัวควบคุมจะถูกควบคุมและการควบคุมจะดำเนินการโดยใช้สาร "สัญญาณ" ในกรณีของเอ็นไซม์ที่เหนี่ยวนำได้ สารนี้คือ "ตัวเหนี่ยวนำ ตัวเหนี่ยวนำ" จะจับกับโมเลกุลของตัวยับยั้งที่ปรับเปลี่ยนรูปร่างเพื่อไม่ให้ปรับตัวเข้ากับ DNA ได้อีกต่อไป ในกรณีนี้ เนื่องจากไม่มีตัวยับยั้งระหว่างโปรโมเตอร์กับยีนที่มีโครงสร้าง , ตัวยับยั้งสามารถสร้างโมเลกุล mRNA และจากโมเลกุลโปรตีนเหล่านี้ สารควบคุมจะกลับมามีการควบคุมอีกครั้งซึ่งจะหยุดการผลิต mRNA ใหม่ ดังนั้น โปรตีนใหม่ ในระบบ beta-galactosidase ตัวเหนี่ยวนำคือแลคโตสหรือสารที่คล้ายคลึงกันนี้มาก อนุพันธ์: พวกเขา จะเข้าร่วมในการยับยั้งการทำงานของมันเพื่อให้สามารถสังเคราะห์เอนไซม์ได้ ในกรณีของเอ็นไซม์ที่กดทับได้ สารที่ทำหน้าที่เป็น "สัญญาณ" จะทำหน้าที่เป็นตัวบีบอัดหลัก: ตัวกดจะทำงานก็ต่อเมื่อรวมกับตัวกดหลักเท่านั้น ในระบบฮิสทิดีนซึ่งเกี่ยวข้องกับเอ็นไซม์หลายสิบชนิด มันคือกรดอะมิโนนี้ รวมกับ tRNA ซึ่งเป็นตัวกดคอร์ ฮิสทิดีน
ปฏิกิริยาอัลโลสเตอริก
ปฏิกิริยาอัลโลสเตอริกซึ่งเกี่ยวข้องกับการหยุดการทำงานของเอนไซม์โดยการเปลี่ยนรูปร่าง ทำให้เกิดวิธีการควบคุมกิจกรรมการเผาผลาญของเซลล์ที่แตกต่างกัน ปฏิกิริยาอัลโลสเตอริกช่วยให้ควบคุมได้แม่นยำกว่าระบบตัวเหนี่ยวนำ-ปราบปรามของโอเปอรอน แต่ไม่บรรลุผลที่เป็นประโยชน์ในการยกเว้นการสังเคราะห์สารที่กำหนดตั้งแต่ขั้นตอนแรก - การผลิต mRNA
ระบบควบคุมใน EUCARIUS
มีข้อเท็จจริงบางประการที่ทำให้เชื่อได้ว่าระบบการควบคุมที่คล้ายกับโอเปอรอนนั้นทำงานและโดดเด่นในหมู่พืชและสัตว์ต่าง ๆ โครโมโซมของสิ่งมีชีวิตเหล่านี้แตกต่างอย่างมากจากโครโมโซมของ E. coli และโปรคาริโอตอื่น ๆ การควบคุมยีนในสิ่งเหล่านี้ เซลล์ต่างกันมาก กลไกของ mitosis คือแต่ละเซลล์ของพืชหรือสัตว์ที่กำหนดมีข้อมูลทั้งหมด
พันธุกรรมที่มีอยู่ในไข่ที่ปฏิสนธิแล้ว ดังนั้น ยีนส่วนใหญ่ในเซลล์พิเศษใดๆ จะยังคงไร้ประสิทธิภาพไปตลอดชีวิตของเซลล์ DNA ในเซลล์เหล่านี้มักเกี่ยวข้องกับโปรตีนเสมอ ดังนั้น จึงเป็นไปได้ที่การยับยั้งยีนในยูคาริโอตจำเป็นต้องอาศัยการเชื่อมโยงนี้อย่างแม่นยำ ระหว่าง DNA และโปรตีน