เส้นทางการเผาผลาญนี้เกี่ยวข้องกับหลายระยะ: คอเลสเตอรอลจะถูกถ่ายโอนจากเนื้อเยื่อส่วนปลายไปยังตับก่อนผ่านระบบน้ำเหลือง จากนั้นจึงผ่านกระแสเลือด ส่วนประกอบหลักที่มีส่วนร่วมในการขนส่งโคเลสเตอรอลแบบย้อนกลับคือ HDL, ABCA1 และ apo A-I
ไม่ใช่ลำไส้หรือตับเซลล์ส่วนปลายไม่สามารถลดคอเลสเตอรอลส่วนเกินได้ ดังนั้น เพื่อรักษาสภาวะสมดุลของเซลล์ การมีอยู่ของกลไกที่ทุ่มเทให้กับการกำจัดคอเลสเตอรอลจึงเป็นสิ่งจำเป็น
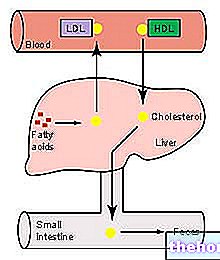
กลไกนี้มุ่งเป้าไปที่การฟื้นฟูตับของคอเลสเตอรอลส่วนเกินที่เรียกว่า "การขนส่งโคเลสเตอรอลย้อนกลับ" (RCT: การขนส่งคอเลสเตอรอลย้อนกลับ).
(ฟอสโฟลิปิดและโคเลสเตอรอล) ซึ่งนำไปสู่การประกอบและสร้างอนุภาค HDL ที่เจริญเต็มที่สารตั้งต้นของ HDL
ขั้นตอนแรกของการขนส่งโคเลสเตอรอลแบบย้อนกลับประกอบด้วยการผลิตโดยลำไส้และตับของสารตั้งต้นของ HDL ดิสคอยด์ซึ่งแสดงอะโพโปรตีนบนผิวของพวกมัน (ส่วนใหญ่เป็น ApoA-I)
ดังนั้น โมเลกุลสารตั้งต้นของ HDL ที่เรียกว่าพรี-B-HDL จะถูกปลดปล่อยออกมา ซึ่งประกอบด้วยคอเลสเตอรอลและไขมันในปริมาณที่น้อยมาก โดยเฉพาะอย่างยิ่ง ฟอสโฟลิปิดการปรากฏตัวของโมเลกุลสารตั้งต้นเหล่านี้ที่ระดับต่อพ่วงช่วยส่งเสริมการถ่ายโอนคอเลสเตอรอลอิสระ (FC) ส่วนเกิน - ที่รั่วออกจากเซลล์เนื้อเยื่อส่วนปลาย - ไปยัง apo AI ผ่านการแทรกแซงของตัวขนส่งเมมเบรนที่เรียกว่า ATP-binding cassette A1 (ABCA1 )
สารขนส่งนี้ถูกแปลเป็นภาษาท้องถิ่นบนผิวเซลล์และในเยื่อหุ้ม Golgi และสามารถขนส่งไขมันจากอุปกรณ์ Golgi ไปยังเยื่อหุ้มเซลล์ ดังนั้นจึงอำนวยความสะดวกในการไหลออก
ณ จุดนี้ ทันทีที่โคเลสเตอรอลอิสระเข้าสู่ HDL ดั้งเดิม เอนไซม์ในพลาสมาที่มีต้นกำเนิดจากตับ เรียกว่าพลาสมาเลซิติน-คอเลสเตอรอล acyltransferase เอนไซม์นี้แปลงโคเลสเตอรอลอิสระที่รวมอยู่ในพรี-B-HDL ให้เป็นเอสเทอร์ของคอเลสเตอรอล โดยเปลี่ยนพรี-บี-เอชดีแอลให้อยู่ในรูปแบบ α-HDL ที่เจริญเต็มที่ ในทางปฏิบัติ การสะสมโคเลสเตอรอลอย่างต่อเนื่องในแกนไลโปโปรตีนจะเปลี่ยน HDL แบบดิสคอยด์ให้เป็นอนุภาคทรงกลมและอวบอ้วน ซึ่งสามารถรับอะโพโปรตีนจากอนุภาคไลโปโปรตีนที่อุดมไปด้วยไตรกลีเซอไรด์และรวมเข้าด้วยกัน
ในกระบวนการทั้งหมด Apolipoprotein AI มีบทบาทสำคัญในการกระตุ้นทั้งกิจกรรมของผู้ขนส่ง ABCA1 และของ LCAT เนื่องจาก ApoAI เป็น apolipoprotein ที่มีตัวแทนมากที่สุดใน HDL ความเข้มข้นในพลาสมาจึงเกี่ยวข้องโดยตรงกับระดับ HDL คอเลสเตอรอล
โปรดทราบ: กระบวนการเอสเทอริฟิเคชันเป็นสิ่งสำคัญในการป้องกันการแพร่กระจายซ้ำของโคเลสเตอรอลจาก HDL ไปยังพลาสมาเมมเบรน กลไกนี้ใช้ประโยชน์จากกรดไขมันในตำแหน่งที่สองที่มีอยู่ในโมเลกุลฟอสฟาติดิลโคลีน
กระบวนการเอสเทอริฟิเคชันที่อาศัย LCAT นั้นจะเปลี่ยนโมเลกุลพรี-B-HDL ให้เป็นทรงกลม α-HDL "ที่โตเต็มที่" ของพวกมัน จากนั้นไลโปโปรตีนเหล่านี้จะถูกส่งไปยังตับ โดยจะปล่อยโคเลสเตอรอลตามเส้นทางที่แตกต่างกันสองทาง
เส้นทางตับแรก
ในกรณีแรก HDL ที่อุดมไปด้วยคอเลสเตอรอล esterified จะถ่ายเทไขมันนี้ไปยังไลโปโปรตีนที่อุดมไปด้วยไตรกลีเซอไรด์ (ไลโปโปรตีนความหนาแน่นต่ำและต่ำมาก) ซึ่งตับจะสกัดกั้นผ่านตัวรับจำเพาะ (LDL-R) และขับออกจากการไหลเวียน
จุดมุ่งหมายคือการส่งคอเลสเตอรอลที่ส่วนปลายไปยังตับผ่านระบบตัวรับ LDL จากนั้น "ปลดปล่อย" HDL จากคอเลสเตอรอลส่วนเกินที่ระดับส่วนปลาย เพื่อให้กลับมาใช้ได้อีกครั้งเพื่อรับจากเนื้อเยื่อ โดยการลดคอเลสเตอรอล HDLs ยอมรับไตรกลีเซอไรด์เพื่อแลกเปลี่ยนและสิ่งนี้เกิดขึ้นด้วยโปรตีนการถ่ายโอนคอเลสเตอรอลเอสเทอร์ (CETP)
หน้าที่ของโปรตีนนี้จึงเป็นประโยชน์ต่อการกระจายและความสมดุลของเอสเทอร์คอเลสเตอรอลและไตรกลีเซอไรด์ระหว่างไลโปโปรตีน HDL, LDL, IDL, VLDL, ไคโลไมครอนและส่วนที่เหลือของไคโลไมครอน นำไปสู่การเสริมคุณค่าในไตรกลีเซอไรด์ของ HDL ด้วยค่าใช้จ่ายของคอเลสเตอรอลเอสเทอร์และการลดขนาดของ HDL
เส้นทางตับที่สอง
วิถีทางที่สองเกี่ยวข้องกับตัวรับ SR-B1 ในตับสำหรับ HDL ที่อุดมไปด้วยเอสเทอริไฟด์โคเลสเตอรอล ในกรณีที่ไม่มีการย่อยสลายส่วนโปรตีนของ HDL ควบคู่ไปด้วย ซึ่งจากนั้นจะถูกนำกลับมาใช้ใหม่ ในทางปฏิบัติ เอนไซม์นี้ช่วยให้ HDL ถูกล้างออกจากเนื้อหาและสร้าง pre-B-HDL ใหม่
อย่างไรก็ตาม ส่วนหนึ่งของ HDL และ ApoA-I นั้นถูกทำให้อยู่ภายในและเสื่อมสภาพที่ระดับ lysosomal ทั้งในเซลล์ตับและไต การดูดซึมที่เป็นสื่อกลางโดย SR-B1 นั้นมีประสิทธิภาพมากขึ้นโดยการทำงานของไลเปสตับ ความสามารถในการสร้าง HDL ใหม่โดยการไฮโดรไลซ์ฟอสโฟลิปิดที่พื้นผิวและทำให้การไหลของเอสเทอริฟายด์โคเลสเตอรอลจากแกนไลโปโปรตีนไปยังพลาสมาเมมเบรน (มีการตั้งสมมติฐานระหว่าง เนื่องจาก ApoE นั้นมีส่วนเกี่ยวข้องกับการคัดเลือกเนื่องจากหนูที่ขาดยีน ApoE จะแสดงประสิทธิภาพของเส้นทางนี้ลดลง) SR-BI ส่วนใหญ่แสดงออกในตับ ต่อมหมวกไต และรังไข่




















-cos-dove-si-trova-e-tossicit.jpg)