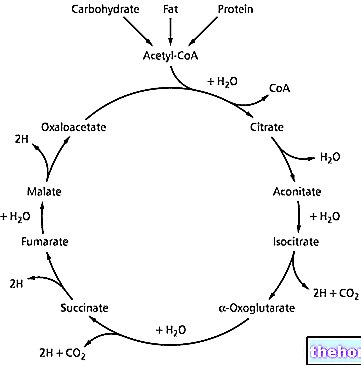
2,3 diphosphoglycerate (2,3 DPG) เป็นสารประกอบที่ได้มาจากผลิตภัณฑ์ขั้นกลางของไกลโคไลซิส มันมีความเข้มข้นโดยเฉพาะอย่างยิ่งในระดับเม็ดเลือดแดงเนื่องจากเซลล์เม็ดเลือดแดง - ปราศจากไมโทคอนเดรีย - ใช้ประโยชน์จากการเผาผลาญแลคตาซิดแบบไม่ใช้ออกซิเจน (การหมักน้ำตาลกลูโคสแบบโฮโมแล็กติก) เพื่อให้ได้พลังงาน

เฮโมโกลบินเป็นโปรตีน tetrameric ซึ่งประกอบด้วยหน่วยย่อยสี่หน่วย สองอัลฟาและสองเบต้า แต่ละหน่วยประกอบด้วยส่วนของโปรตีน (โกลบิน) และ heme (กลุ่มเทียมที่ผูกมัดออกซิเจน) 2,3-diphosphoglycerate ยึดติดกับสาย Beta ที่กระชับและลดความสัมพันธ์ของเฮโมโกลบินสำหรับออกซิเจน
การผูกมัดของ 2,3 DPG กับเฮโมโกลบินเกิดขึ้นเมื่อฮีโมโกลบินอยู่ในรูปแบบ deoxygenated ในขณะที่ถูกละลายในปอดโดยการจับฮีโมโกลบินกับออกซิเจน อันที่จริง เมื่อฮีโมโกลบินไปถึงเนื้อเยื่อ β เชน เป็นคนแรกที่ปล่อยออกซิเจนและการสูญเสียนี้เกี่ยวข้องกับการแทนที่ของโมโนเมอร์จากศูนย์ ทันทีที่ช่อง hydrophilic ภายในเปิดขึ้น DPG จะเข้าและจับกับ tetramer เพื่อสร้างพันธะเฮเทอโรโพลาร์ระหว่างกลุ่มที่มีประจุลบกับไลซีนและฮิสทิดีน สารตกค้างของสายโซ่ เบต้า มีประจุบวก โครงสร้างที่เสถียรยังสามารถปล่อยออกซิเจนของสายโซ่ α สองสายได้ อย่างไรก็ตามในปอดจะมีกระบวนการย้อนกลับ ที่ความดันออกซิเจนสูง โซ่ α จะผูกมัดกับสายโซ่แรก และ DPG จะถูก "บีบ" และขับออกจากเตตระเมอร์ ซึ่งช่วยให้เกิดพันธะของสายโซ่ออกซิเจน-β ได้ง่ายขึ้น
2,3 bisphosphoglycerate ไม่สามารถจับกับเฮโมโกลบินของทารกในครรภ์ได้เนื่องจากโมเลกุลนี้ไม่มีสายโซ่ B ซึ่ง 2,3 DPG จับ สิ่งนี้อธิบายความสัมพันธ์ที่ใกล้ชิดกับออกซิเจนของเฮโมโกลบินในครรภ์มากกว่าเฮโมโกลบินของมารดาซึ่งเป็นลักษณะเฉพาะที่ช่วยให้เลือดของทารกในครรภ์ สกัดออกซิเจนจากเลือดของมารดา